Golgi Complex: Terminologi, forekomst, Morfologi, Struktur og andre detaljer
Golgi-kompleks: Terminologi, forekomst, Morfologi, Struktur, Strukturel og Biokemisk Polarisering, Kemisk Sammensætning, Funktioner og Origins!
I 1898 opdagede Golgi en retikulær struktur i cytoplasma ved hjælp af en sølvfarvningsmetode. Navnet "Golgi-apparat", der generelt gives til denne struktur, er forvirrende, fordi det antyder et konkret forhold til cellens fysiologiske processer.
I dag synes det mere hensigtsmæssigt at bruge navnet "Golgi substans" eller "Golgi kompleks" for at henvise til dette materiale, der har særlige farvende egenskaber. Fordi dets brydningsindeks svarer til matrixens, er Golgi-komplekset vanskeligt at observere i levende celler. Anvendelsen af elektronmikroskopet har givet et tydeligt billede af denne komponent, og dens submikroskopiske struktur er blevet afsløret.
I årevis blev Golgi-komplekset antaget at være en artefakt af forskellige fiksering og farvningsprocedurer. Med andre ord troede mange videnskabsmænd, at strukturen, der blev observeret under mange mikroskopiske procedurer og betegnes Golgi, faktisk ikke eksisterede i den levende celle. Guilliermond (1923), Parat (1927), Walker og A lien (1921) opstod der tvivl med hensyn til eksistensen af Golgi-komplekset.
Terminologi af Golgi kompleks:
Holmgren refererede til Golgi-komplekset som trophospongium (Cajal betegner det som Golgi-Holmgren kanalerne). Baker brugte udtrykket lipokondrier på grund af det formodede lipidindhold. Udtrykket Dalton Complex blev givet efter navnet på dets observatør Dalton i 1952. Sjostrand foreslog termen cytomembraner til Golgi-systemet.
Sosa har foreslået følgende nomenklatur for Golgi-komplekset:
1. Golgiokinesis :
Opdeling af Golgi-apparatet under atomafdeling.
2. Golgiosomer :
Korpuser produceret af Golgiogenese kaldes som Golgiosomer, der beskrives som Golgi materiale i hvirvelløse dyr.
3. Golgiolyse :
Opløsningsproces for Golgi-apparatet.
4. Gogiorrhexis :
Fragmentering på Golgi-apparatet.
5. Golegiogenese :
Formation og differentiering af Golgi krop under embryonisk udvikling.
6. Golgio-cytoarkitektur :
Studie af cellens struktur i forhold til Golgi-apparatet.
Hændelse:
Golgi-komplekset forekommer i alle celler undtagen de prokaryote celler (nemlig Mycoplasma, bakterier og blågrønalger) og eukaryote celler af visse svampe, sædceller af bryophytter og pteridofytter, celler af modne sigtør af planter og modne sæd og rødt blod dyr af dyr.
Golgi-kompleksets morfologi:
Golgi-kompleksets morfologi varierer fra celle til celle afhængig af typen af celle, hvori de findes. To former for Golgi-kompleks er blevet observeret.

1. Lokaliseret form :
I polariserede celler af hvirveldyr (som har base og apex), forekommer Golgi-komplekset enkeltvis og indtager en fast position. Det ligger mellem kernen og sekretorisk stang. Dette kan ses bedst i skjoldbruskkirtelceller, i eksokrine celler i bugspytkirtlen og tarmslimhinden.
2. Diffuseret form :
I nogle specialiserede celler af hvirveldyr (nerveceller og leverceller) findes i flere planteceller og i celler af hvirvelløse dyr flere enheder af Golgi-komplekset spredt sammen med elementerne i endoplasmatisk retikulum. Hver enhed hedder et diktyosom. I leverceller forekommer der op til 50 diktyosomer pr. Celle, og i visse planteceller kan deres antal nå op til hundreder.
Form :
Formen på Golgi-komplekset er ret variabel i forskellige somatiske celletyper af dyr. Selv i samme celle er der variationer i forskellige funktionelle trin. Formen er dog konstant med hver celletype. Det varierer i form fra en kompakt masse til et spredt trådformet netværk.
Nummer:
Antallet af Golgi-stabler pr. Celle varierer enormt afhængigt af celletypen - fra så få som en til hundrederne. Der er en enkelt stor i nogle celler, mens der i Paramoeba er to. I Stereomyxa (en Amoeba-art) er der mange Golgi-komplekser. Nerveceller, leverceller og de fleste planteceller har også flere Golgi-komplekser, hvor der er ca. 50 i leverceller.
I algerorizoider ses mere end 25.000 Golgi-komplekser (Sievers 1965). Golgi-komplekset kan endda tegne sig for en stor del af cellevolumenet i nogle specialiserede celler. Et eksempel er tarmcellen i tarmepitelet, som udskiller slim i tarmen; glycoproteinerne i slim glycosyleres hovedsageligt i Golgi-komplekset.
Størrelse:
Størrelsen er ligeledes variabel. Det er stort i nerve- og kirtelceller og små i muskelcellerne. Generelt er Golgi-komplekset veludviklet, mens cellen er i aktiv tilstand. Når cellen bliver gammel, formindsker komplekset gradvist i størrelse og forsvinder.
Position:
Placeringen af Golgi-komplekset er relativt fast for hver celletype. I cellerne med ektopodermal oprindelse polariseres Golgi-komplekset fra tidspunktet for embryonal tilstand mellem kernen og periferien (Cajal, 1914). I de sekretoriske eksokrine celler, der generelt har en typisk polarisation, findes Golgi-komplekset mellem kernen og sekretorisk polen.
I de endokrine kirtler er polariteten af denne organoid variabel, undtagen i skjoldbruskkirtlen, hvor den er orienteret mod midten af follikelet. I de yngre celler og ofte i de ældre ligger det mest almindeligt på den ene side af kernen, men i visse tilfælde kan det helt omgiver det. I muskelganglioniske celler er positionen perinuclear.
Detaljeret struktur af Golgi kompleks:
Dalton og Felix (1954) beskrev Golgi-komplekset i råtters epididymier efter at have taget de første elektronmikrografier.
Den følgende beskrivelse af Golgi-komplekset er en sammensat baseret på flere forfatteres arbejde:
1. Cisternae :
Cisternae eller saccules svarer til den glatte overflade ER, og vises i snit som stakke af tæt adskilte membran afgrænsede sacs. Antallet af saccula varierer fra 4 til 8 i de fleste dyre- og plantecelletyper. I Euglena kan tallet gå op til 20.
Scculernes membran er ca. 60 til 70 A ° i tykkelse, som omslutter et hulrum ca. 150 A ° bredt, hvis kanter ofte dilateres. Ifølge de fleste forfattere er der to veldefinerede ansigter af cisternae, dvs. konvekse og konkave; sidstnævnte betegnes generelt som det modne eller formende eller distale overflade, og den konvekse side antages at være den umodne eller excitere eller proximale overflade, idet den cisterane ligning i parallel array adskilles fra hinanden med et mellemrum på ca. 200 til 300 A 0 .
Hvad der holder dem sammen er endnu ikke kendt, men i få celler ses et tyndt lag af elektronopakket, undertiden tæt materiale mellem de saccula, som i visse regioner er mere fremtrædende, som Amos og Grimstone (1968) anvendte begrebet noder. Mollenhauer et al., (1973) udforskede i nogle detaljer sammenhængende elementer og plaques i visse plante Golgi komplekse.

2. Tubuler :
Fra det perifere område af cisternae opstår et komplekst anastomoserende fladt netværk af rør med en diameter på 300 til 500 A °. Clowes and Juniper (1969) har sammenlignet dette rørformede netværk med blonder af blonder.
3. Vesikler :
Vesiklerne er små dråbelignende sacs, som forbliver fastgjort til tubuli ved cisternaens periferi.
De er af følgende to typer:
(a) glatte vesikler :
De glatte vesikler har en diameter på 20 til 80 μm. De indeholder sekretorisk materiale (så ofte kaldes sekretoriske vesikler) og er bøjet ud fra enderne af cisternal tubuli inden for nettet. Ofte mere end en tubuleforbindelse og formodentlig at fylde en enkelt formende vesikel.
(b) overtrukne vesikler :
De belagte vesikler er sfæriske fremspring, omkring 50 μm i diameter og med en grov overflade. De findes i periferien af organellen, sædvanligvis i enderne af enkeltrør og er morfologisk helt forskellige fra de sekretoriske vesikler. Deres funktion er ukendt.
4. Golgian vacuoles :
Disse er store afrundede sacs til stede på Golgi modning. Disse dannes enten ved den udvidede cisternae eller ved sammensmeltningen af sekretoriske vesikler. Vakuolerne er fyldt med noget amorft eller granulært stof.
Golgi-komplekset er strukturelt og biokemisk polariseret:
Golgi-komplekset har to adskilte ansigter: en cis, eller formende ansigt og et trans, eller modent ansigt. Cis-overfladen er tæt forbundet med en glat overgangsdel af det grove ER. I sekretoriske celler findes transfaget ansigt tættest på plasmamembranen. Her findes de store sekretoriske vesikler udelukkende i forbindelse med overfladen af en Golgi-stabel, og membranen af en dannende sekretorisk vesikel er ofte kontinuert med den af trans-ansigtet af den sidste ("trans-mest") cisterna.
I modsætning hertil er de små Golgi-vesikler lokaliseret mere jævnt langs stakken, og proteiner er almindeligt tænkt at komme ind i en Golgi-stak fra ER på cis-siden og for at afslutte flere destinationer på trans-siden; Men hverken deres nøjagtige vej gennem Golgi-komplekset eller hvordan de rejser fra cisterna til cisterna langs hver stak er kendt.
Golgi-kompleksets to flader er biokemisk forskellige. For eksempel kan en variation i tykkelsen af Golgi-membranerne påvises over stakken i visse tilfælde, hvor de på cis-siden er tyndere (ER-lignende), og de på trans-siden er tykkere (plasmamembran-lignende).
Mere slående er de resultater, der opnås, når visse histokemiske tests anvendes i forbindelse med elektronmikroskopi for at lokalisere bestemte proteiner inden for Golgi-komplekset. Nogle af disse tests afslører membranbundet enzym, aktiviteter der viser en distinkt polaritet i deres lokalisering inden for Golgi-stakken.
Et særligt spændende biokemisk fund var opdagelsen af, at lysosomale enzymer, såsom sur phosphatase, er koncentreret med Golgi-stakkens trans-cisterne og i nogle af de belagte vesikler i nærheden. Dette antyder, at specifikke vesikler, der forlader lysosomer, samles i denne region.
Sekretoriske proteiner findes ved histokemiske metoder i alle de stablede cisternae, selvom de store sekretoriske vesikler, hvori disse produkter er koncentreret og kun er associeret med de mest Golgi-cisterna.
Kemisk sammensætning:
Hvad angår Golgi-kompleksets kemiske sammensætning er det blevet påvist, at følgende stoffer er til stede:
1. Fosfolipider :
Fosfolipidsammensætning af Golgi-membraner er mellemliggende mellem dem af endoplasmiske membraner og plasmamembraner.
2. Proteiner og enzymer :
Golgi-komplekset fra forskellige plante- og dyreceller viser store variationer i protein- og enzymindholdet. Nogle af enzymerne er ADPase, ATPase, NADPH cytochrom-C-reduktase, glycosyltransferaser, galactosyltransferase, thiaminpyrophosphat etc.
3. Kulhydrater :
Både plante- og dyreceller har nogle almindelige kulhydratkomponenter, såsom glucosarin, galactose, glucose, mannose og fructose. Plant Golgi mangler sialinsyre, men forekommer i høj koncentration i rotterlever. Nogle kulhydrater som xylulose og arabinose findes kun i planteceller.
4. Vitamin С :
Den del af vitamin С, der er lagret i Golgi-komplekset, er blevet vist af Tomitte. Ifølge ham lagrer Golgi-komplekset vitamin С og frigiver det langsomt ind i cytoplasmaet i tilstrækkelig grad til forebyggelse af oxidation af celleprodukterne.

Golgi-kompleksets funktioner:
1. Formation af Acrosom under Spermiogenese :
Under modningen af sæd spiller Golgi-komplekset en rolle i dannelsen af acrosom (Burgos og Fawcett, 1955).

I tidlige stadier vises Golgi som en sfærisk krop, der består af cisternae arrangeret i parallelle stakke og talrige små vesikler. Den senere klæbte altid fra cisternae. Efterhånden som udviklingen fortsætter, bliver Golgi-komplekset uregelmæssigt i form, og store vakuloler dannes ved udvidelser af cisternal sacs.
I midten af disse store vakuole eller vakuoler er der en tæt granul, det proacrosomale granulat. Dette granulat, der er afledt af Golgi-komplekset, fortsætter med at vokse i vakuolen ved en fremgangsmåde kendt som accretion. Denne vacuole og granulat nærmer sig den forreste pol af den nukleare membran, der udgør acrosomal granulat.
Ved forlængelsen af spermatidet spredes den acrosomale vesikel over den nukleare overflade og falder til sidst sammen med nukleærmembranen og danner capmaterialet. Det acrosomale granul bliver akrosomet, som ligger ved kernens apex og tilsyneladende omfatter visse enzymer involveret i befrugtningsprocessen.
2. Syntese og sekretion af polysaccharider:
Undersøgelser af bægerceller ved autoradiografi og elektronmikroskopi har etableret sammenhængen mellem proteinsyntese, kulhydrattilsætning og sulfatering. Tarmcellerne i tyktarmen producerer mucigen. Dette sekretoriske materiale indeholder en stor del af kulhydratet.
Golgi-komplekset findes lige over kernen. Mod den frie overflade af cellen forstørres gradvist mucigengranulerne. Den nærliggende cisternae af Golgi-komplekset viser ingen hævelse, men på en afstand over stakken bliver de distale cisternae helt pludselig omdannet til slimhindegranuler. Den distale cisternae konverteres løbende til mucigen granulat hver 2-4 minutter. Ny proximal cisternae dannes i kompensation.
3. Rolle i sekretion:
Golgi-komplekset anses for at spille en rolle i sekretorisk funktion af en celle. Men spørgsmålet er, at de udskiller eller syntetiserer nogle stoffer selv, eller de er simpelthen et butikhus, hvor de sekretoriske produkter, der udskilles et andet sted i cellen, simpelthen opbevares og koncentreres.


Fra studierne af Palade et al. I 1962 er denne sekretoriske cyklus nu veldefineret og omfatter fire trin i tilfælde af akutceller i bugspytkirtlen og de er:
(i) Indarbejdelsen af aminosyrer i protein på overfladen af groft endoplasmatisk retikulum.
(ii) Overførsel af disse fremspringende sekretoriske proteiner i cisternae af groft endoplasmatisk retikulum.
(iii) Den intracellulære transport af disse proteiner til Golgi-komplekset.
(iv) Migrering af zymogengranuler mod cellens apex, hvor de udledes i lumen.
4. Rolgi kroppens rolle i oogenesis :
Srivastava (1965) har givet en kort gennemgang af Golgi-komplekset under oogenese. Ifølge Afzelius (1956) består Golgi-komplekset af et hav-urchin æg, som ses under elektronmikroskop, af stabler af lameller, der danner vægge af flade poser, der lejlighedsvis kan blive opsvulmet.
Der er nogle indikationer på tværgående opdelinger af disse organer. Sotelo (1959) og Sotelo og Porter (1959) har beskrevet Golgi-komplekset i rotte-ovum som set under elektronmikroskopet og fundet juxtra nuklear lokalisering af denne organelle i tidlige oocytter.
I den næste fase løser disse i fragmenter, og i tredje fase bevæger disse sig mod cortexen. I alle disse tilfælde forbliver deres struktur af tæt pakket arrays af slanke, dobbelte profiler (flade sager) og sfæriske vesikler.
I de tidlige oocytter er komplekset kompakt organiseret. I senere stadier findes diskrete bundler af profiler, omgivet af små vesikler, spredt i den kortikale zone. I de tidlige oocytter er Golgi-komplekset og centrosomet tæt forbundet.
5. Absorption af forbindelser :
Hirsch et al., Har opdaget, at når jernsukker bliver fodret til et dyr, bliver jern absorberet på Golgi-komplekset (Kedrowsky). Van Teel har vist, at Golgi-systemer også absorberer forbindelser af kobber og guld. Kedrowsky har vist, at Golgi-komplekset af Opalina kan absorbere bismutose (sammensætning af albumin og bismuth) og protargol (sammensætning af albumin og sølv). Således angiver Kirkman og Severinghaus, at Golgi-komplekset fungerer som en kondensationsmembran til koncentration af produkter i dråber eller granuler.
6. Plante cellevæg dannelse :
Plantenes cellevæg består af fibriller, der overvejende indeholder polysaccharider sammen med nogle lipider og proteiner. Under cytokines dannes en celleplade mellem de to datterkerner og har omkring den en membran, som senere bliver dattercellernes plasmamembran. Der er tydelige tegn på, at polysacchariderne dannes i Golgi-komplekset og overføres til den nye cellevæg, der lægges ned, mens cellerne stadig vokser.

Stoffer som pectiner og hemicelluloser, som danner matrixen af cellepladen, der adskiller plasmamembranerne, bidrager også af Golgi-komplekset.
7. Formation af intracellulære krystaller :
I den marine isopod er Limnoria lingmorum, som er en burrowing form, der er til stede midglands, hvis celler består af krystaller. Disse rækker op til 30 A ° i længde og 15 A ° tyk. Det er blevet bevist, at disse krystaller er dannet af Golgi-kompleks og er kendt for at indeholde protein og jern. De er uden at lukke membranen og sædvanligvis sfærisk i form. De er bekymrede for den sekretoriske aktivitet.
8. Dampdannelse af mælkeprotein :
I den lakterende brystkirtlen hos mus fremstilles proteindråber, der er relateret til Golgi-komplekset. Disse dråber åbner sædvanligvis på celleoverfladen ved fusion af deres omgivende membran med plasmamembranen.
9. Fremstilling af lysosomer og vakuoler :
Primære lysosomer udformes fra Golgi-membranerne på samme måde som de sekretoriske vesikler. Der er gode tegn på, at dictyosomer akkumulerer hydrolytiske enzymer i deres mere modne regioner. Nogle vacuoler i planteceller har vist sig at indeholde små mængder hydrolytiske enzymer, og disse formodes at være blevet afledt af Golgi-komplekset.
10. Pigmentdannelse:
I mange mammale tumor- og cancerceller er Golgi-komplekset blevet beskrevet som oprindelsesstedet for pigmentgranulater (melanin).
11. Regulering af væskebalance :
En homologi er blevet foreslået mellem Golgi-komplekset og den kontraktile vakuol af lavere Metazoa og Protozoa. Den kontraktile vakuol udviser overskydende vand fra cellen. I visse Protozoer er Golgi-komplekset også bekymret for regulering af væskebalance.
Oprindelse af Golgi Complex:
Der er foreslået tre forskellige kilder, hvorfra der kan opstå et nyt Golgi-kompleks:
1. Fra endoplasmatisk retikulum :
Essner og Novik®ff (1962) og Beams and Kessel (1968) har foreslået, at Golgi-cisternaen stammer fra ER. Det grove endoplasmatiske retikulum efter syntetisering af specifikke proteiner mister ribosomer og ændres til glat ER. Små forbigående vesikler klemmer af fra glat ER. Disse overgår til diktyosom.
Ved at nå diktyosoms formningsflade danner disse sikringer en ny cisternae og bidrager dermed til dens vækst. Ved sammensmeltningen af disse vesikler formes nye cisternae kontinuerligt på formningsfladen og på den modneside nedbryder de gamle cisternae i sekretoriske vesikler. Således udviser Golgi et fænomen med membranstrøm. '
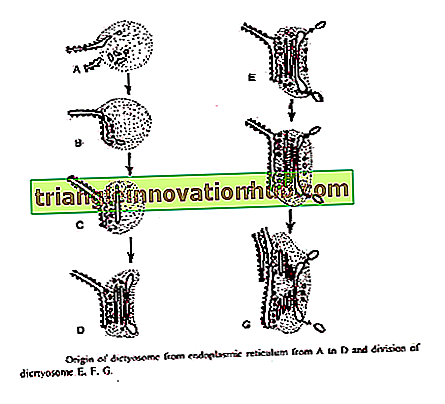

Bouch (1965) beskrev Golgis oprindelse fra ydre membran af nuklear kuvert i brune alger. Vesiklerne klemmes ud fra ydre nuklearmembran, som smelter sammen til dannelse af cisternae på diktyosoms formningsflade.
Tilstedeværelsen af udelukkelseszoner i forbindelse med glat ER eller nuklearmembran, forekomsten af udelukkelseszoner i sovende frø af højere planter og dannelsen af dictyosom fra disse zoner i spirende frø giver beviser til støtte for de ovennævnte to teorier om oprindelsen af dictyosome.
3. Ved opdeling af eksisterende diktyosom :
Det er blevet observeret, at i løbet af celledeling i både planter og dyr øges antal diktyosomer, og antallet af diktyosomer i hver dattercelle lige efter division er næsten lig med antallet i modercellen forud for opdeling, fra dåser og andre direkte observationer på de opdelte celler er det antaget, at dictyosomer også opdeles under celledeling.