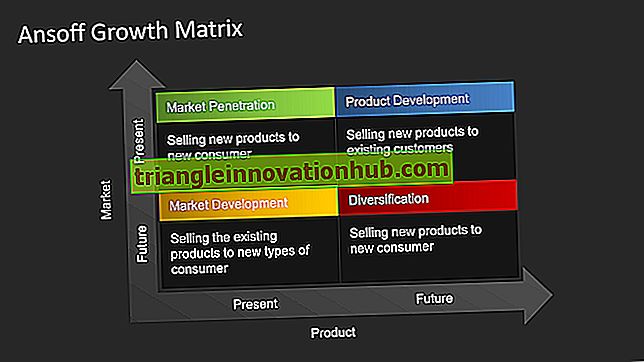
5 Faktorer der påvirker genetisk ligevægt
Nogle af de væsentligste faktorer, der påvirker den genetiske ligevægt og fremkalder variabiliteten i befolkningen, er som følger: (A) Mutationer (B) Rekombinationer under genoptræning (C) Genetisk drift (D) genmigration (genstrømning) .
Ifølge Hardy-Weinberg Equilibrium-loven forbliver den relative frekvens af alleler i befolkningen konstant fra generation til generation i en population af seksuelt reproducerende organismer, når:
(i) Befolkningen er stor nok til at undgå stikprøver
(ii) Parring foregår tilfældigt
(iii) Mutation finder ikke sted, eller hvis det gør, er satsen samme i begge retninger
(iv) Alle befolkningens medlemmer overlever og har samme reproduktionshastighed
Faktorer der påvirker Hardy-Weinberg-ligevægten:
Der er fem faktorer, som påvirker den genetiske ligevægt og inducerer variabiliteten i befolkningen. Disse faktorer kaldes evolutionære agenter.
(A) Mutationer:
Disse er kendetegnet ved:
(i) Disse er pludselige, store og arvelige ændringer i det genetiske materiale.
(ii) Mutationer er tilfældige (ikke-diskriminerende) og forekommer i alle retninger.
(iii) De fleste mutationer er skadelige eller neutrale. Det anslås, at kun en ud af 1.000 mutationer er nyttige.
(iv) Mutationsmængden er meget lav, dvs. en pr. million eller en pr. million million genie loci. Men mutationshastigheden er tilstrækkelig til at producere betydelig genetisk variabilitet.
(v) Visse mutationer er præadaptive og forekommer selv uden eksponering for et bestemt miljø. Disse udtrykker og bliver kun fordelagtige, når de er udsat for nye omgivelser, som kun vælger de præadaptive mutationer, der tidligere fandt sted.
Eksistensen af præadaptive mutationer i Escherichia coli blev eksperimentelt demonstreret af Esther Lederberg (1952) i replikplateringsforsøg (Forklaret i Neo-Darwinisme).
(vi) På basis af mængden af genetisk materiale involveret er mutationer af tre typer (tabel 7.15):

(vii) På basis af deres oprindelse er mutationer af to typer (tabel 7.16):
Tabel 7.16. Forskelle mellem spontane og inducerede mutationer.
tegn | Spontane mutationer | Inducerede mutationer |
1. forårsaget af 2. Frekvens af mutationer 3. årsager | Af naturlige midler kaldes også naturlige mutationer eller baggrundsmutationer. Meget lavt (ca. en million gen eller mere). Ikke sikkert, mange cellulære produkter, f.eks. Formaldehyd, salpetersyre, peroxider osv. Virker som mutagener. | Af mand Hurtigere Visse fysiske {f.eks. Strålingstemperaturer osv.) Og kemiske midler kaldes mutagener. |
(viii) Betydningen af mutationer:
(a) Mutationer skaber og opretholder variationer i en befolkning.
(b) Disse introducerer også nye gener og alleler i en genpool (figur 7.45).

(c) Akkumulering af mutationer over en række generationer kan føre til sammensætning.
(B) Rekombinationer under seksuel reproduktion:
Rekombination involverer omfordeling af gener af kromosomer. Chancer for rekombination er mere i de organismer, der gennemgår seksuel reproduktion, der involverer gametogenese efterfulgt af befrugtning.
Seksuel reproduktion indebærer rekombinationer i tre faser:
(i) Krydsning over (figur 7.46):
Det indebærer udveksling af genetisk materiale mellem ikke-søsterkromatiderne af homologe kromosomer.
Overgangsmekanisme involverer følgende faser:

(a) Synapser involverer parring af homologe kromosomer under zygotenstadiet af Prophase 1 af meioser til dannelse af bivalenter.
(b) Tetraddannelse som hver bivalent er dannet af fire kromatider under pachytenstadiet af meiose I.
(c) Chiasma dannelse på grund af coiling af ikke-søster kromatider af homologe kromosomer til dannelse af X-formede punkter, kaldet chiasma.
d) Krydsning indebærer udveksling af genetisk materiale.
(ii) Ved uafhængigt udvalg af kromosomer:
Under metafase-I arrangerer bivalenterne ved spindelens ækvator i to ækvatoriale eller metafaseplader. Under anafase I bevæger homologe kromosomer mod modsatte poler. Dette kaldes disjunction og resulterer i reduktion af kromosom nummer. Variationer forekommer under chancenordningen af bivalenter under metafase I af meiose.
Antal producerede rekombinationer afhænger af antallet af bivalenter i organismen og er givet ved formlen 2 n (hvor n repræsenterer antallet af bivalenter), fx i mennesket, vil antallet af mulige kombinationer af gameterne være 8, 6 x 106 (2 23 ).
(iii) Ved tilfældig befrugtning:
Det involverer chancen fusion af gameter, fx i mennesket er antallet af mulige typer zygote dannet 70 x 10 12 . Det er sådan som enhver sæd med enhver kombination af gener kan smelte sammen med noget æg med en hvilken som helst kombination af gener.
Betydning:
På grund af rekombination er det kun omfordeling af allerede eksisterende tegn, og der produceres ingen nye gener, men det fører til omfordeling af forskellige træk til forskellige individer i en befolkning. Forskellige kombinationer bringer mangfoldighed i genotype og fænotype af forskellige organismer. Så rekombination er et middel for evolution.
(C) genetisk drift:
Det er den tilfældige ændring i frekvensen af alleler, der forekommer tilfældigt udsving. Det er kendetegnet ved:
(i) Det er en binomial prøveudtagningsfejl i genpoolen, dvs. at alleler, som danner genpuljen i den næste generation, er en prøve af allelerne af den nuværende befolkning.
(ii) Genetisk drift påvirker altid frekvenser af alleler og er omvendt proportional med befolkningens størrelse. Så genetisk drift er vigtigst i meget små populationer, hvor der er øgede chancer for indavl, som øger hyppigheden af individer homozygote til recessive alleler, hvoraf mange måske er skadelige.
(iii) Genetisk drift sker, når en lille gruppe adskiller sig fra en større befolkning og måske ikke har alle allellerne eller kan afvige fra forældrenes befolkning i frekvenserne af visse gener. Dette forklarer for forskellen mellem øpopulationer og fastlandsbefolkningen.
(iv) I en lille befolkning kan en chance begivenhed (fx snevejr) øge hyppigheden af et tegn med lille adaptiv værdi.
(v) Genetisk drift kan også fungere gennem grundlæggervirkning. Her kan genetisk drift forårsage dramatiske ændringer i allelfrekvenserne i en population afledt af små grupper af kolonisatorer, kaldet grundlæggere, til et nyt habitat.
Disse grundlæggere har ikke alle de alleler, der findes i deres kildepopulation. Disse grundlæggere bliver hurtigt forskellige fra forældrenes befolkning og kan danne en ny art, fx udvikling af Darwin-finches på Galapagosøerne, som sandsynligvis var afledt af nogle få grundlæggere.
(vi) Befolkning flaskehals (Fig. 7.47):
Det er en reduktion i allelfrekvenser forårsaget af en drastisk reduktion i befolkningsstørrelsen, der kaldes befolkningskrasj, fx fald i fangstpopulationen i Afrika på grund af overjagt. Da den givne genpool er begrænset, forhindrer populationsflaskehalsen ofte arten til at genoprette sin tidligere rigdom, så ny befolkning har en meget begrænset genpool end den større forældrepopulation.

(D) genmigration (genstrømning):
De fleste populationer er kun delvis isolerede fra andre populationer af samme art. Normalt forekommer nogle migrationsemigrationer (flytning ud af nogle individer ud af en befolkning) eller indvandring (indførsel af nogle medlemmer af en population til en anden population af samme art) mellem befolkningerne.
Indvandring resulterer i tilføjelse af nye alleler i den eksisterende genpool og ændrer allelfrekvenserne. Graden af ændringer i allel frekvenser afhænger af forskellene mellem genotyper af indvandrere og indfødte befolkning.
Hvis der ikke er mange genetiske forskelle, vil indlæsningen af et lille antal indvandrere ikke ændre allelfrekvenserne meget. Men hvis befolkningen er genetisk helt anderledes, kan en lille mængde indvandring medføre store ændringer i allel frekvenser.
Hvis de migrerende individer blandes med medlemmerne af lokalbefolkningen, kaldet hybridisering, kan disse bringe mange nye alleler ind i værtspopulationens lokale genpool. Dette kaldes gen migration. Hvis de interspecifikke hybrider er frugtbare, kan disse indlede en ny udviklingstendens, der fører til dannelse af nye arter.
Denne tilsætning eller fjernelse af alleler, når individer går ind eller forlader en population fra en anden lokalitet, hedder genstrøm. Ubegrænset genflow mindsker forskellene mellem genpoolerne og reducerer særpræget mellem forskellige populationer.
Tabel 7.17. Forskelle mellem mikro-evolution og makro-evolution.
tegn | Mikro-evolution | Makro-evolution |
1. Ændringsniveau 2. Udviklerens art 3. Eksempler | På det genetiske niveau. Sort eller art. Oprindelse af forskellige sorter af afgrødeplanter og tamdyr. | På jorden skala af geologisk tid. Højere kategorier som Genus, Familie, Ordre, Klasse eller Filum, også kaldet kvante evolution. Evolution af forskellige pattedyrtyper fra de fælles forfædre primitive insektivorer ved adaptiv stråling. |
(E) Naturligt udvalg:
(i) Definition:
Den proces, hvormed forholdsvis bedre tilpassede individer ud af en heterogen befolkning er begunstiget af naturen over de mindre tilpassede individer hedder naturligt valg.
ii) mekanisme:
Processen med naturlig udvælgelse virker gennem differentiel reproduktion.
Det betyder, at de personer, som bedst er tilpasset miljøet, overlever længere og reproducerer i højere grad og producerer flere afkom end dem, der er mindre tilpassede.
Så formgiverne bidrager forholdsmæssigt til større procentdel af gener til genpoolen i næste generation, mens mindre tilpassede individer producerer færre afkom. Hvis differentiel reproduktion fortsætter i et antal generationer, vil generne af de personer, der producerer flere afkom, blive overvejende i populationens genpulje (figur 7.48):

På grund af seksuel kommunikation er der fri bevægelse af gener, således at den genetiske variabilitet, der forekommer hos visse individer, gradvist spredes fra den ene dem til den anden, fra dem til befolkning og derefter på nabostillede søsterpopulationer og endelig på de fleste medlemmer af en arter. Så naturlig udvælgelse forårsager progressive ændringer i genfrekvenser, dvs. frekvensen af adaptive gener øges, mens frekvensen af mindre adaptive gener falder.
Så naturligt udvalg af neo-darwinisme fungerer som en kreativ kraft og opererer gennem komparativ reproduktiv succes. Akkumulering af sådanne variationer fører til oprindelsen af en ny art.
(iii) Typer af naturlig udvælgelse:
De tre forskellige typer af naturlige selekteringer er:
1. Stabilisering eller afvejning af udvælgelse:
Det fører til eliminering af organismer med overspecialiserede tegn og opretholder en homogen population, der er genetisk konstant. Det favoriserer de gennemsnitlige eller normale fænotyper, samtidig med at de udelukker personer med ekstreme udtryk. I dette opnår flere personer gennemsnitlig karakterværdi.
Det reducerer variationen, men ændrer ikke middelværdien. Det resulterer i meget langsom udviklingstid. Hvis vi tegner en grafisk kurve af befolkningen, er den klokkeformet. Den klokkeformede kurve indsnævres på grund af eliminering af ekstreme varianter (figur 7.49).

Eksempel:
Sickle-celleanæmi hos mennesker (Forklaret i Neo-Darwinisme).
2. Retnings- eller Progressiv udvælgelse:
I dette udvalg ændres befolkningen til en bestemt retning sammen med miljøændringer. Da miljøet gennemgår en kontinuerlig ændring, overlever organismerne, der har erhvervet nye tegn, og andre elimineres gradvist (figur 7.50).

I dette elimineres individer i den ene ekstrem (mindre tilpasset), mens individer i andre ekstremer (mere tilpassede) er begunstiget. Dette skaber flere og flere tilpassede individer i befolkningen, når et sådant valg fungerer i mange generationer. I denne type udvælgelse erhverver flere personer værdi end den gennemsnitlige karakterværdi.
Eksempler:
Industrielle melanisme (Forklaret i neo-darwinisme):
I dette faldt antallet af de lysfarvede møller (Biston betularia) gradvist, mens den af melaniske møller (B. carbonaria) steg med retningsvalg.
DDT-resistente myg (Forklaret i Neo-Darwinisme):
I dette blev følsomme myg elimineret og resistente dem steg i antal. Så øget befolkningen af modstandsdygtige myg viser retningsmæssig udvælgelse.
3. Disruptive selection:
Det er en slags naturlig udvælgelse, der favoriserer ekstreme udtryk for bestemte træk for at øge variansen i en befolkning. Det bryder en homogen befolkning i mange adaptive former. Det resulterer i afbalanceret polymorfisme.
I denne type udvælgelse erhverver flere personer perifert karakterværdi i begge ender af fordelingskurven. Denne form for udvælgelse er sjælden og eliminerer de fleste medlemmer med gennemsnitlig ekspression, så der produceres to toppe i fordelingen af et træk (figur 7.51).

Eksempel:
I havet er de tre typer snegle dvs. hvidfarvet; brunfarvet og sortfarvet er til stede. De hvide farvede snegle er usynlige, når de er dækket af havkirtler. De sorte farvede snegle er usynlige, når sten er bar. Men brune farvede snegle bliver spist af rovdyr under begge forhold. Så disse elimineres gradvist.