Kvælstofmetabolisme i planter: Måder, udnyttelse og andre detaljer
Kvælstofmetabolisme i planter: Måder, udnyttelse og andre detaljer!
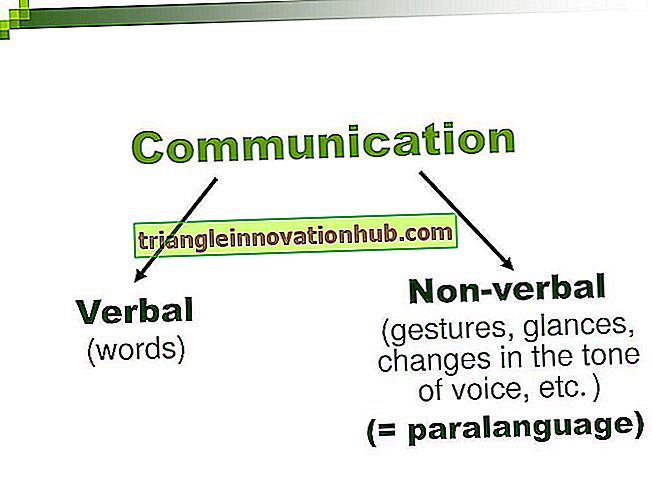
Der er et stort udvalg af nitrogenholdige forbindelser i en celle. Protein, som udgør den væsentlige bestanddel af protoplasma, nukleinsyrer, der bestemmer de morfologiske og fysiologiske grænser for en art såvel som de enkelte medlemmer deraf, enzymer, vitaminer, hormoner, alkaloider, glycosider osv. Er alle nitrogenholdige.
Atmosfæren er den ultimative nitrogenkilde. Ud over det lille antal arter, der kan fastsætte det atmosfæriske nitrogen og udnytte det til at syntetisere de utallige kvælstofstoffer, de har brug for, opnår alle planter deres kvælstofbehov fra jorden.
Nitraterne og de ammoniakholdige salte, der udgør en del af jordens mineralmiljø, optages af rødderne og anvendes i kvælstofmetabolisme af planten. Planterne opnår således deres kvælstofbehov på fire forskellige måder:
(1) som molekylært nitrogen;
(2) som organisk nitrogen;
(3) som ammoniak nitrogen og
(4) som nitratkvælstof.
Udnyttelse af molekylært kvælstof:
Kun få få organismer er i stand til kvælstoffiksering, dvs. udnyttelse af atmosfærisk nitrogen i syntesen af nitrogenholdige forbindelser. Den bedst kendte nitrogenbindende symbiotiske bakterie er Rliizobium. Denne bakterie lever i jorden for at danne rodknuder i planter, der tilhører familien Leguminosae, såsom bønner, gram, jordnød og sojabønne.

Nogle frie levende bakterier som Azotobacter og Clostridium pasteurianum er også i stand til at fastsætte atmosfærisk nitrogen. Rhodospirillum, Rhodopseudomonas og Chromatium, som kan fotosyntesere, kan også rette nitrogen. Men reduktionen af kvælstof sker gennem en reduceret elektrondonor, såsom sulfid, svovl, hydrogen eller organiske forbindelser, og ikke ved hjælp af lysenergi.
Andre kvælstoffastgørelsesorganismer er nogle af de blågrønne alger, såsom Anabaena og Nostoc. Normalt er de frit levende, men nogle gange danner de symbiotiske foreninger, når de findes i koralloidrødderne af Cycas og blade af vandlevnen Azolla.
Knudeformationen i bælgplanterne er blevet undersøgt i detaljer. Når en del af rodknuden undersøges, forekommer den lyserød på grund af tilstedeværelsen af et pigment kaldet leghemoglobin. Som hæmoglobin er leghemoglobin en iltforskyder. Synthesen af leghemoglobin kræver kobolt, fordi det er en væsentlig del af vitamin B 12, en forbindelse der er nødvendig til fremstilling af pigmentet.
Også molybdæn er afgørende, men dets rolle er ikke kendt for sikker; måske spiller den rollen som en cofactor og letter redox reaktionerne, hvilket fører til reduktion af nitrogen til ammoniak. Det enzym, som katalyserer fikseringen af nitrogen (nitrogenase) funktioner under aerobe forhold. Leghemoglobin kombinerer med ilt og beskytter ilt og beskytter kvælstof.
De nøjagtige reaktioner involveret i reduktionen af molekylært kvælstof til ammoniak er ikke kendt for visse. Omdannelsen af N2 til NH3 er imidlertid ikke en ret proces. De mulige veje i reduktion af molekylært nitrogen til ammoniak er angivet nedenfor.
Udnyttelse af organisk nitrogen:
Mange planter anvender organiske nitrogenforbindelser som aminosyrer og urinstof. Brugen af urinstof ved planter er ikke grundigt udarbejdet, men urease, enzymet, der virker på urinstof, er sandsynligvis hydrolyseret til to ammoniakmolekyler og et af carbondioxid. Ammonien metaboliseres derefter på den sædvanlige måde.
Ammoniumkalisk nitrogen:
Ammoniumsalte anvendes ofte som en kilde til nitrogen ved flere planter. Men beredskaben med hvilken en plante optager en ammoniakforbindelse afhænger af jordens natur og plantens alder. Den ammoniakale radikal absorberet af en plante er indbygget i amider, aminosyrer og proteiner.
Nitrat kvælstof:
Jordbunden indeholder gode mængder nitrater, og de fleste planter får deres kvælstofbehov i den form. Faktisk foretrækker flere planter nitratkvælstof til ammoniakkvælstof. Nitratkvælstofet reduceres til ammoniakform før inkorporering i en af nitrogenforbindelserne i cellen. Allerede i 1895 foreslog Schulze følgende vej til denne reduktion.
Nitrat -> Nitrit -> Hyponitrit -> Hydroxylamin -> Ammoniak
1. Reduktion af nitrat til nitrit:
Det udføres af agenturen af enzyminitratreduktase. Enzymet har FAD som sin protetiske gruppe og bruger NADPH eller NADH som kilde til reduktion af effekt.

Molybdæn er også afgørende for reduktionen af nitrater. Dette fremgår af den kendsgerning, at nitrater i molybdæn - mangelfulde planter ophobes; det tjener som en elektronbærer. Ordningen med elektronoverførsel, der finder sted under reduktionen af nitrater til nitrit, kan afbildes som angivet nedenfor.

2. Nitritreduktion:
Venecko og Verner (1955) viste, at reduktionen af nitrat-nitrogen til aminokvælstof forekommer i intakte blade både i lys og mørke. Tidligere i 1953 viste Evans og Nason at i planteekstrakter, der indeholder NADPH, forsvinder nitrater langsomt, men det ser ud til, at reduktionskraften herfor er tilvejebragt ved fotokemisk opdeling af vand.
2HNO2 + 2H20 -> 2NH3 + 3O2
Et nitritreducerende enzym er endnu ikke isoleret, selvom Nason, Abraham og Averback (1954) forberedte et sådant enzymsystem fra Neurospora. Dette system har vist sig at være i stand til at anvende NADH eller NADPH som hydrogen donor.
Reduktion af hydroxylamin:
Nason (1955) og Frear and Burrel (1955) isolerede enzymet, der omdanner hydroxylamin til ammoniak fra henholdsvis Neurospora og højere planter. Dette enzym er også mistænkt for at være et metalloflavoprotein med mangan som metalkofaktoren. Det anvender også NADH som hydrogen donor.
NH20H + NADH + H + -> NH3 + NAD + H20
Sammenfatning af ligningen i henhold til Schulzes plan, vi har

Til sidst er ammoniak indbygget i aminosyre, enten glutaminsyre eller asparaginsyre. Disse tjener til at indlede en lang række transaminationsreaktioner og derfra til at bygge proteiner.
Aminosyre Metabolisme:
Syntese af aminosyrer forekommer hovedsagelig i rødder og blade, hvor nitrater reduceres. Cellerne i disse regioner skal tilvejebringe de nødvendige organiske syrer, der hovedsagelig dannes i Krebs-cyklus. Som Krebs-cyklus forekommer i mitokondrier, antages det, at lokaliteterne af aminosyresyntese er mitokondrier. Aminosyrer dannes ved hjælp af følgende metoder:
1. Reduktiv Aminering:
I nærværelse af en dehydrogenase og en reducerende effekt (enten NADH2 eller NADPH.) Kan ammoniak kombinere direkte med en keto-syre til dannelse af en aminosyre. Substratet med reduktiv aminering er generelt a-ketoglutursyre, et mellemprodukt med Krebs-cyklus. Det giver anledning til Glutaminsyre. Glutaminsyre anerkendes som indgangsport for uorganisk nitrogen i den metaboliske vej.
Dannelsen af glutaminsyre finder sted gennem to trin. Første a-ketoglutarsyre amineres til a-iminoglutaric, og derefter dannes glutaminsyre gennem mediation af NADPH og glutaminsdehydrogenase.

Ud over a-ketoglutarsyre er andre organiske syrer, som undergår reduktiv aminering, oxaloacetat og pyrodruesyre.
2. Transaminering:
Når en aminosyre er organiseret, danner den udgangspunktet for produktionen af andre aminosyrer. Dette sker gennem transaminationsreaktioner. En transamineringsreaktion består i overføringen af en aminogruppe fra en aminosyre til en organisk syre, således at en anden aminosyre dannes.
En typisk transamineringsreaktion kan være repræsenteret som følger:


Wilson, King og Burris (1954) viste transamineringsreaktioner, der involverede glutaminsyre og sytten andre aminosyrer. Nogle af dem er:
Glutaminsyre + oxaloeddikesyre a-ketoglutarsyre + asparaginsyre
Glutaminsyre 4 Pyruvinsyre a-Ketoglutarsyre + Alanin.
Et vigtigt aspekt ved aminosyre-metabolisme er, at de hverken opbevares eller udskilles.